Hyperlipidemia is one of the conditions that constitute metabolic disorder and it is a common public health problem. The condition is characterized by increased levels of cholesterol, triglycerides and/or lipoproteins; it is a recognized as a risk factor for the onset of many diseases such as type 2 diabetes, non-alcoholic fatty liver disease, and cardiovascular disease. Up to now, the primary drugs for treating hyperlipidemia are statins and monoclonal antibody drugs against proprotein convertase subtilisin/kexin type 9 (PCSK9). The main limitation of statins for long-term use is intolerable side effects. Evolocumab and Alirocumab, two monoclonal antibodies against PCSK9, can effectively decrease the level of low-density lipoprotein cholesterol (LDL-C) in patients with statin intolerance and familial hypercholesterolemia, while causing fewer side effects. However, due to its short half-life and high costs, these monoclonal antibody treatments might result in patients’ non-compliance with medication and considerable economic burden on patients. Given that RNA plays a key role in gene regulation, RNA-based therapeutics have become powerful blueprints for designing new anti-hyperlipidemia drugs. Here, we summarized RNA-based therapeutic strategies and the current clinical trials for RNA drugs in hyperlipidemia treatment.
- Open Access
- Review
From Bench to Bedside: Current Developments in RNA-Based Therapies for Treatment of Hyperlipidemia
- Yufei Zhou,
- Chen Chen *
Author Information
Received: 07 Oct 2022 | Accepted: 06 Nov 2022 | Published: 21 Dec 2022
Abstract
Graphical Abstract
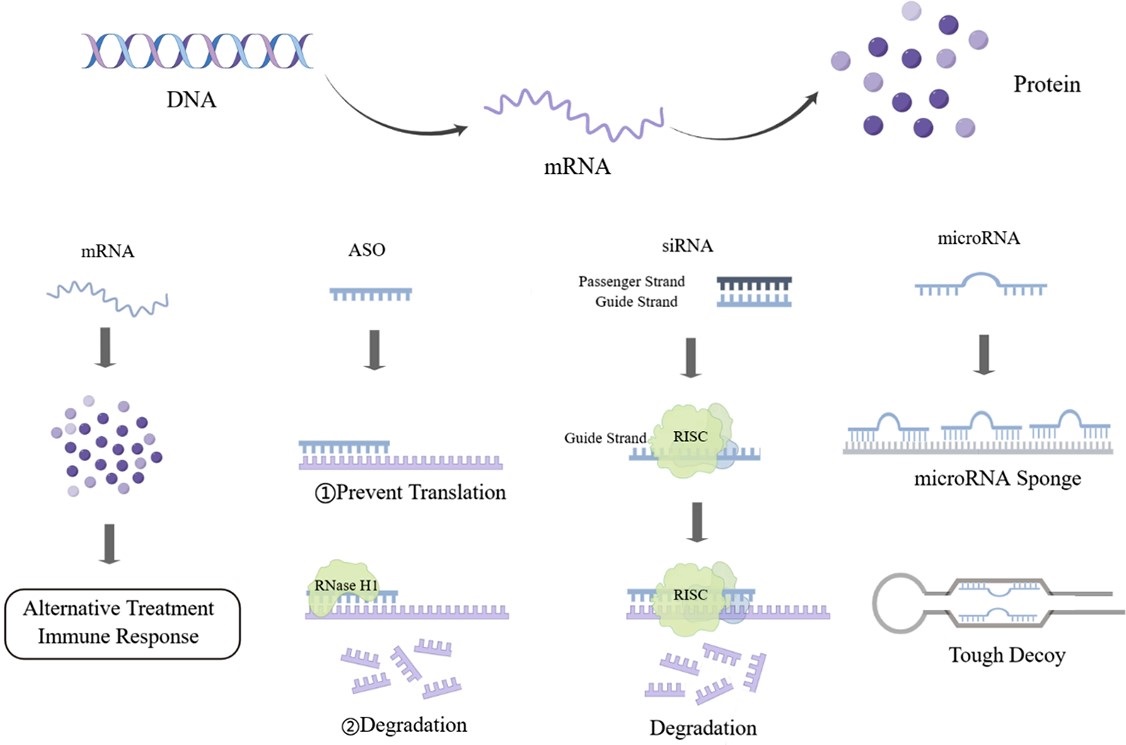
Keywords
hyperlipidemia | microRNA | therapeutic strategies | clinical trial
References
- 1.Rosenson R.S.; Najera S.D.; Hegele R.A. Heterozygous familial hypercholesterolemia presenting as chylomicronemia syndrome. J. Clin. Lipidol., 2017, 11(1): 294-296.
- 2.Averbukh L.D.; Turshudzhyan A.; Wu D.C.; et al. Statin-induced liver injury patterns: a clinical review. J Clin Transl Hepatol, 2022, 10(3): 543-552.
- 3.Vinci P.; Panizon E.; Tosoni L.M.; et al. Statin-associated myopathy: emphasis on mechanisms and targeted therapy. Int. J. Mol. Sci., 2021, 22(21): 11687.
- 4.Carmena R.; Betteridge D.J. Diabetogenic action of statins: mechanisms. Curr. Atheroscler. Rep., 2019, 21(6): 23.
- 5.Galicia-Garcia U.; Jebari S.; Larrea-Sebal A.; et al. Statin treatment-induced development of type 2 diabetes: from clinical evidence to mechanistic insights. Int. J. Mol. Sci., 2020, 21(13): 4725.
- 6.Santos R.D.; Stein E.A.; Hovingh G.K.; et al. Long-term evolocumab in patients with familial hypercholesterolemia. J. Am. Coll. Cardiol., 2020, 75(6): 565-574.
- 7.Santos R.D.; Ruzza A.; Hovingh G.K.; et al. Evolocumab in pediatric heterozygous familial hypercholesterolemia. N. Engl. J. Med., 2020, 383(14): 1317-1327.
- 8.Blom D.J.; Harada-Shiba M.; Rubba P.; et al. Efficacy and safety of alirocumab in adults with homozygous familial hypercholesterolemia: the ODYSSEY HoFH trial. J. Am. Coll. Cardiol., 2020, 76(2): 131-142.
- 9.Nissen S.E.; Stroes E.; Dent-Acosta R.E.; et al. Efficacy and tolerability of evolocumab vs ezetimibe in patients with muscle-related statin intolerance: the GAUSS-3 randomized clinical trial. JAMA, 2016, 315(15): 1580-1590.
- 10.Hu B.; Zhong L.P.; Weng Y.H.; et al. Therapeutic siRNA: state of the art. Signal Transduction Targeted Ther., 2020, 5(1): 101.
- 11.Tsimikas S. RNA-targeted therapeutics for lipid disorders. Curr. Opin. Lipidol., 2018, 29(6): 459-466.
- 12.Kazi D.S.; Penko J.; Coxson P.G.; et al. Updated cost-effectiveness analysis of PCSK9 inhibitors based on the results of the FOURIER trial. JAMA, 2017, 318(8): 748-750.
- 13.Yu A.M.; Choi Y.H.; Tu M.J. RNA drugs and RNA targets for small molecules: principles, progress, and challenges. Pharmacol. Rev., 2020, 72(4): 862-898.
- 14.Qin S.G.; Tang X.S.; Chen Y.T.; et al. mRNA-based therapeutics: powerful and versatile tools to combat diseases. Signal Transduction Targeted Ther., 2022, 7(1): 166.
- 15.Katzmann J.L.; Packard C.J.; Chapman M.J.; et al. Targeting RNA with antisense oligonucleotides and small interfering RNA: JACC state-of-the-art review. J. Am. Coll. Cardiol., 2020, 76(5): 563-579.
- 16.Kulkarni J.A.; Witzigmann D.; Thomson S.B.; et al. The current landscape of nucleic acid therapeutics. Nat. Nanotechnol., 2021, 16(6): 630-643.
- 17.Wada F.; Yamamoto T.; Ueda T.; et al. Cholesterol-GalNAc dual conjugation strategy for reducing renal distribution of antisense oligonucleotides. Nucleic Acid Ther., 2018, 28(1): 50-57.
- 18.Kubczak M.; Michlewska S.; Bryszewska M.; et al. Nanoparticles for local delivery of siRNA in lung therapy. Adv. Drug Delivery Rev., 2021, 179: 114038.
- 19.Weng Y.H.; Li C.H.; Yang T.R.; et al. The challenge and prospect of mRNA therapeutics landscape. Biotechnol. Adv., 2020, 40: 107534.
- 20.Hu B.; Li B.; Li K.; et al. Thermostable ionizable lipid-like nanoparticle (iLAND) for RNAi treatment of hyperlipidemia. Sci. Adv., 2022, 8(7): eabm1418.
- 21.Ammirati E.; Veronese G.; Brambatti M.; et al. Fulminant versus acute nonfulminant myocarditis in patients with left ventricular systolic dysfunction. J. Am. Coll. Cardiol., 2019, 74(3): 299-311.
- 22.Soh J.; Iqbal J.; Queiroz J.; et al. MicroRNA-30c reduces hyperlipidemia and atherosclerosis in mice by decreasing lipid synthesis and lipoprotein secretion. Nat. Med., 2013, 19(7): 892-900.
- 23.Zhang T.P.; Shi H.T.; Liu N.N.; et al. Activation of microRNA-378a-3p biogenesis promotes hepatic secretion of VLDL and hyperlipidemia by modulating ApoB100-Sortilin1 axis. Theranostics, 2020, 10(9): 3952-3966.
- 24.Ng R.; Wu H.; Xiao H.; et al. Inhibition of microRNA-24 expression in liver prevents hepatic lipid accumulation and hyperlipidemia. Hepatology, 2014, 60(2): 554-564.
- 25.Bernardo B.C.; Gregorevic P.; Ritchie R.H.; et al. Generation of MicroRNA-34 sponges and tough decoys for the heart: developments and challenges. Front. Pharmacol., 2018, 9: 1090.
- 26.Li H.P.; Fan J.H.; Zhao Y.R.; et al. Nuclear miR-320 mediates diabetes-induced cardiac dysfunction by activating transcription of fatty acid metabolic genes to cause lipotoxicity in the heart. Circ. Res., 2019, 125(12): 1106-1120.
- 27.Jeong D.; Yoo J.; Lee P.; et al. miR-25 tough decoy enhances cardiac function in heart failure. Mol. Ther., 2018, 26(3): 718-729.
- 28.Zheng C.Y.; Khoo C.; Furtado J.; et al. Apolipoprotein C-Ⅲ and the metabolic basis for hypertriglyceridemia and the dense low-density lipoprotein phenotype. Circulation, 2010, 121(15): 1722-1734.
- 29.Witztum J.L.; Gaudet D.; Freedman S.D.; et al. Volanesorsen and triglyceride levels in familial chylomicronemia syndrome. N. Engl. J. Med., 2019, 381(6): 531-542.
- 30.Hegele R.A. Apolipoprotein C-Ⅲ inhibition to lower triglycerides: one ring to rule them all?. Eur. Heart J., 2022, 43(14): 1413-1415.
- 31.Graham M.J.; Lee R.G.; Brandt T.A.; et al. Cardiovascular and metabolic effects of ANGPTL3 antisense oligonucleotides. N. Engl. J. Med., 2017, 377(3): 222-232.
- 32.Horton J.D.; Cohen J.C.; Hobbs H.H. Molecular biology of PCSK9: its role in LDL metabolism. Trends Biochem. Sci., 2007, 32(2): 71-77.
- 33.Collaboration NCDRF. Repositioning of the global epicentre of non-optimal cholesterol. Nature, 2020, 582(7810): 73-77.
- 34.Blanco-Domínguez R.; Sánchez-Díaz R.; De La Fuente H.; et al. A novel circulating MicroRNA for the detection of acute myocarditis. N. Engl. J. Med., 2021, 384(21): 2014-2027.

How to Cite
Zhou, Y.; Chen, C. From Bench to Bedside: Current Developments in RNA-Based Therapies for Treatment of Hyperlipidemia. International Journal of Drug Discovery and Pharmacology 2022, 1 (1), 7. https://doi.org/10.53941/ijddp.v1i1.141.
RIS
BibTex
Copyright & License

Yufei Zhou, Chen Chen
This work is licensed under a Creative Commons Attribution 4.0 International License.
Contents
References
 Suite 4002 Level 4, 447 Collins Street, Melbourne, Victoria 3000, Australia
Suite 4002 Level 4, 447 Collins Street, Melbourne, Victoria 3000, Australia General Inquiries: info@sciltp.com
General Inquiries: info@sciltp.com